A collection of manuscripts published by the Group since 2007. Clicking each title reveals the reference, a short description, and an image associated with that manuscript. Graduate student names in bold font.
Experience-Dependent Plasticity During Auditory Learning (2018)
Ross MT, Flores D, Bertram R, Johnson F, Wu W, Hyson RL. Experience-Dependent Intrinsic Plasticity Auditory Learning. J Neurosci. 2018 Dec 12. pii: 1036-18. doi: 10.1523/JNEUROSCI.1036-18.2018. PubMed PMID: 30541908 PubMed Link.
It is well established that learning involves plasticity of the synapses between neurons. However, the activity of a neural circuit can also be dramatically altered by changes in the intrinsic properties (ion channels) of the component neurons. Matthew Ross (Graduate Student, Hyson Lab) conducted a series of experiments demonstrating experience-dependent changes in the intrinsic physiology of neurons in the cortical premotor nucleus HVC in juvenile zebra finches during auditory learning of a tutor's song. Tutor deprivation does not "arrest" development of intrinsic properties, but rather, results in neurons with a premature adult-like physiological phenotype (see example below). Diana Flores (Graduate Student, Bertram Lab) then developed biophysical models to identify the ion channel types modulated by experience. It is possible that auditory learning involves a form of non-synaptic plasticity and that experience-dependent suppression of specific ion channels may work in concert with synaptic plasticity to promote vocal learning.
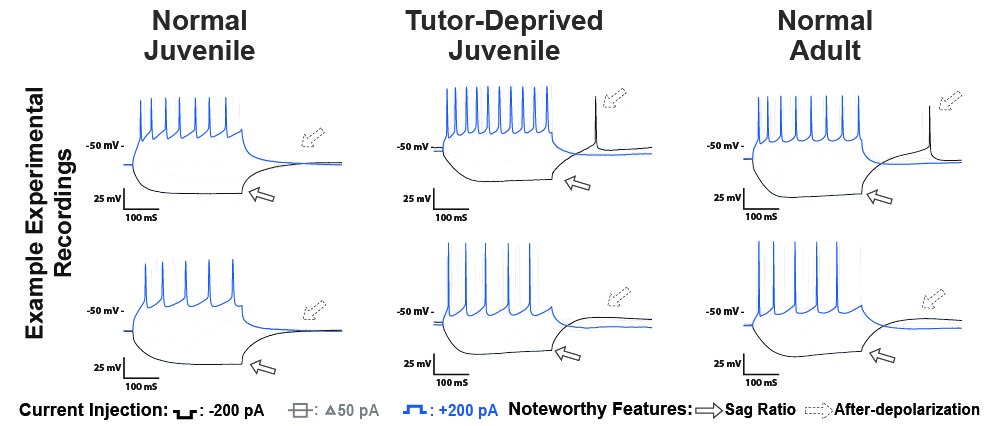
Female Zebra Finches Do Not Sing Yet Share Neural Pathways Necessary For Singing In Males (2018)
Shaughnessy DW, Hyson RL, Bertram R, Wu W, Johnson F. Female Zebra Finches Do Not Sing Yet Share Neural Pathways Necessary For Singing In Males. J Comp Neurol. 2018 Oct 29. doi: 10.1002/cne.24569. PubMed PMID: 30370534 PubMed Link.
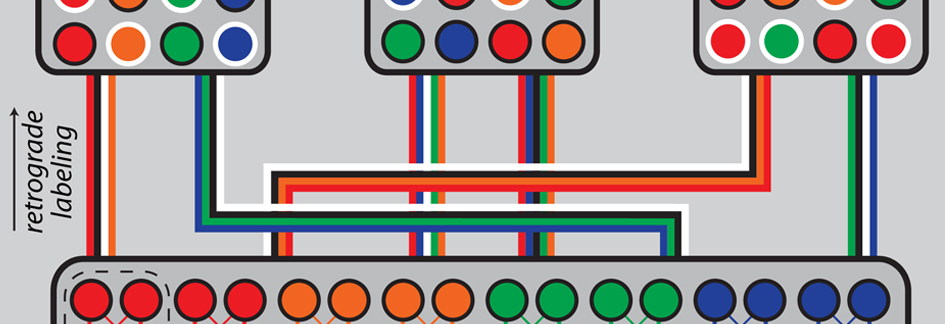
Adult female zebra finches, which do not produce learned songs, have long been thought to possess only vestiges of the forebrain network that supports learned song in males. This view ostensibly explains why females do not sing - many of the neural populations and pathways that make up the male song control network appear rudimentary or even missing in females. For example, classic studies of vocal-premotor cortex (HVC, acronym is name) in male zebra finches identified prominent efferent pathways from HVC to vocal-motor cortex (RA, robust nucleus of the arcopallium) and from HVC to the avian basal ganglia (Area X). In females, by comparison, the efferent targets of HVC were thought to be only partially innervated by HVC axons (RA) or absent (Area X). Here, using a novel visually guided surgical approach to target tracer injections with precision (see image below), Derrick Shaughnessy (Graduate Student, Johnson Lab) mapped the extrinsic connectivity of the adult female HVC for the first time. He found that female HVC shows a mostly male-typical pattern of afferent and efferent connectivity, including robust HVC innervation of RA and Area X. As noted by earlier investigators, Derrick found large sex differences in the volume of many regions that control male singing (male > female). However, sex differences in volume were diminished in regions that convey ascending afferent input to HVC. The findings do not support a vestigial interpretation of the song control network in females. Instead, Derrick's findings support the emerging view that the song control network may have an altogether different function in nonsinging females.
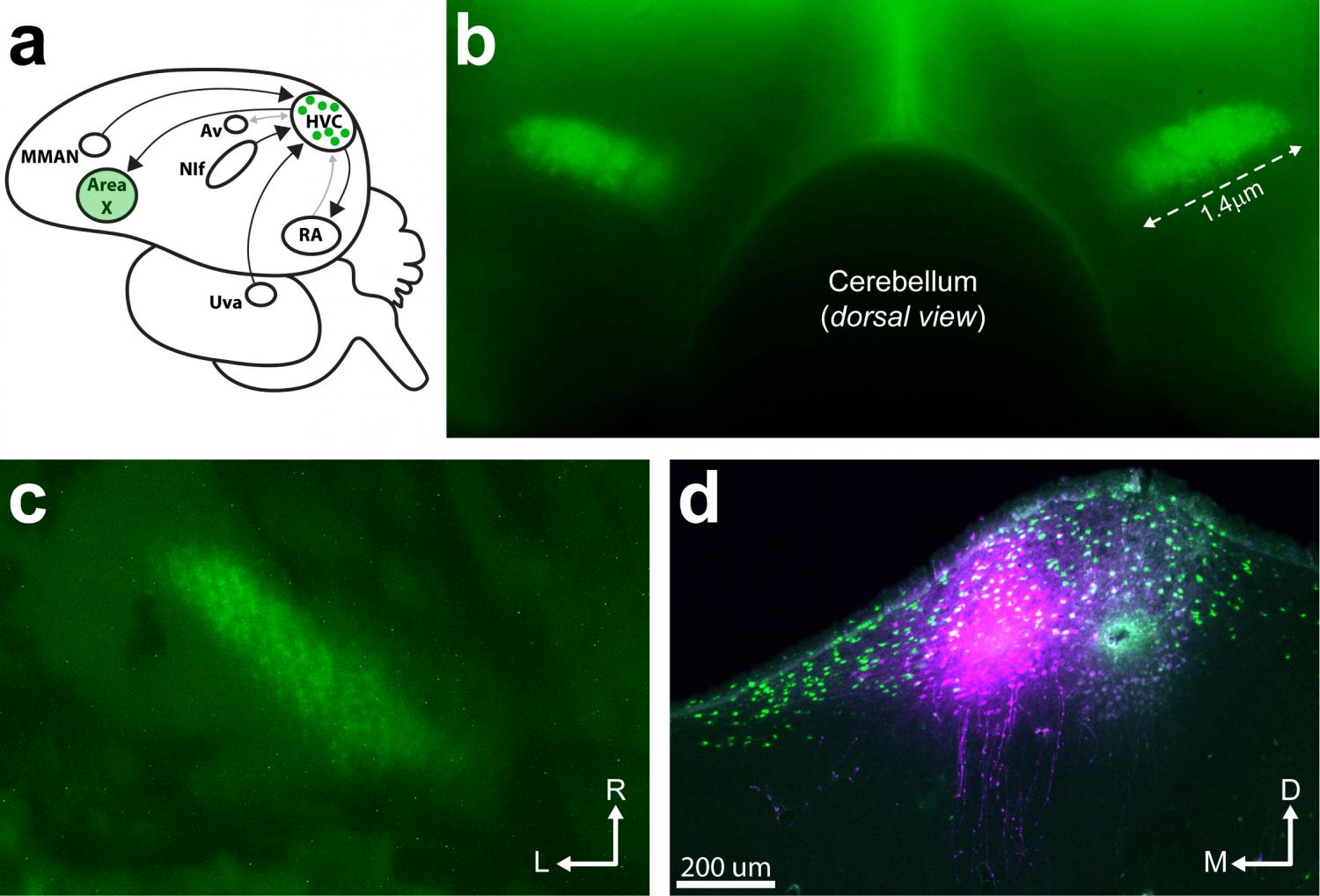
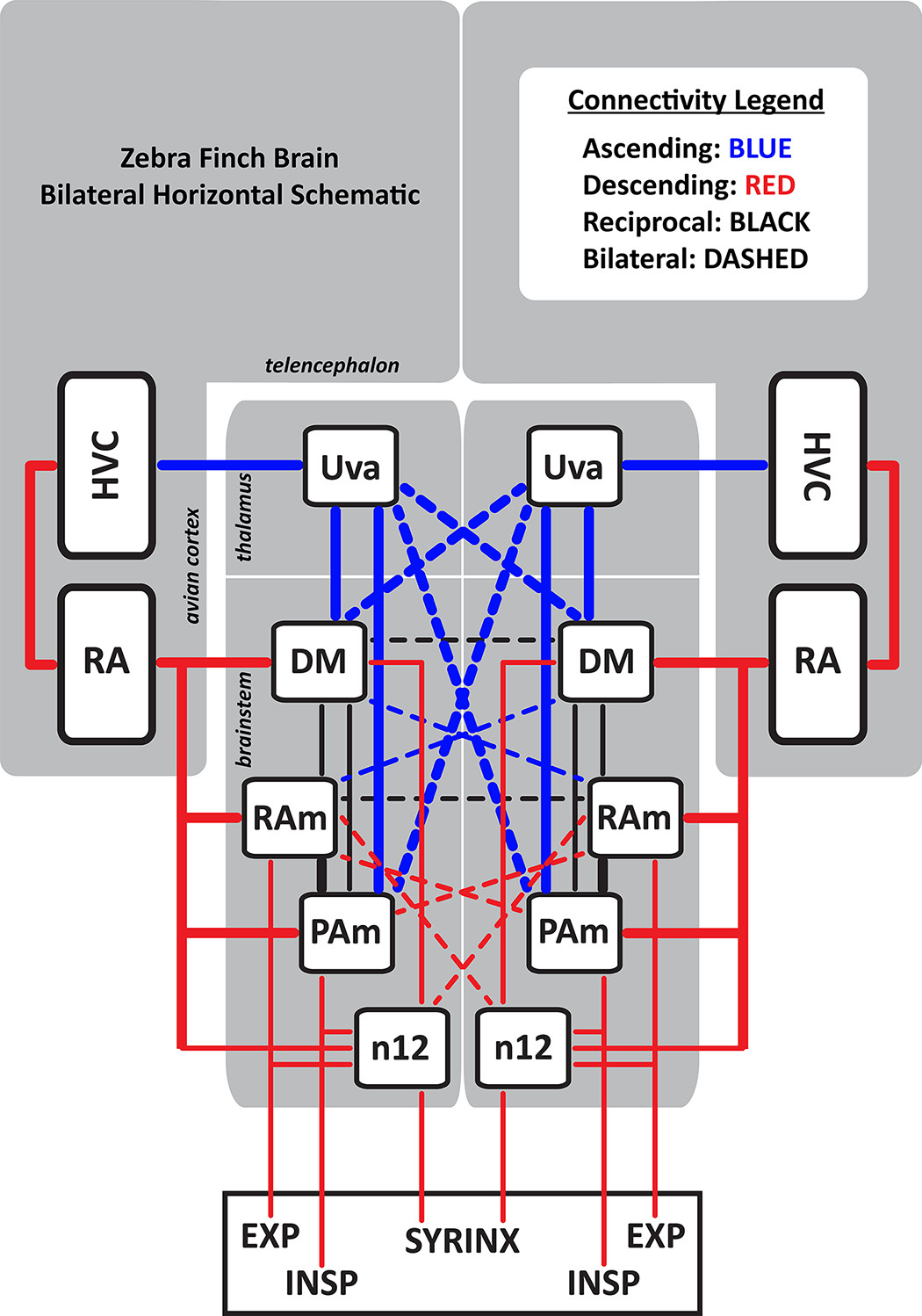
Interhemispheric Dominance Switching in a Neural Network Model for Birdsong (2018)
Galvis D, Wu W, Hyson RL, Johnson F, Bertram R. Interhemispheric dominance switching in a neural network model for birdsong. J Neurophysiol. 2018 Sep 1;120(3):1186-1197. doi: 10.1152/jn.00153.2018. Epub 2018 Jun 20. PubMed PMID: 29924715 PubMed Link.
With various elements of the learned song encoded across both hemispheres, how is activity between the two hemispheres coordinated during the production of song? This is a particularly tricky problem for the avian brain, which lacks a corpus callosum. The image below depicts the dense brainstem and thalamic connectivity where communication between the two hemispheres would be possible. Based on this pattern of connectivity, Danny Galvis (Graduate Student, Bertram Lab) took the lead on development of a computational network model of interhemispheric coordination that accommodates the neural activity patterns that drive singing (and of course, breathing). Importantly, the model also captures the disruptive behavioral effects caused by unilateral cooling and electrical stimulation of the song network. The model makes testable predictions that will be used to validate the model framework and explain the effects of other perturbations of the network, such as unilateral ablation of ascending or descending pathways.
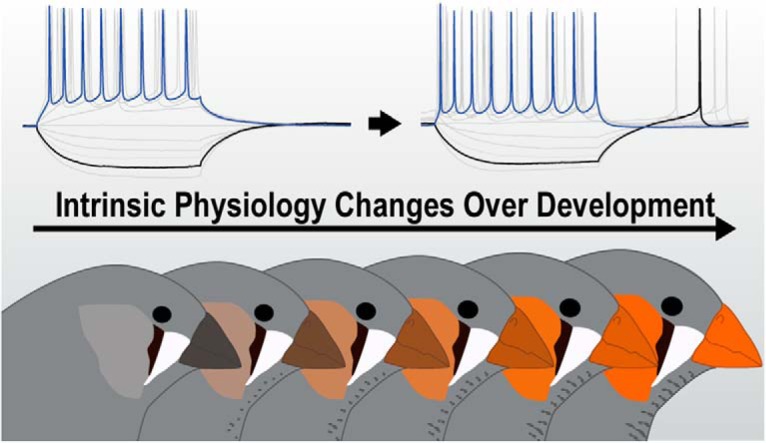
Neuronal Intrinsic Physiology Changes During Development of a Learned Behavior (2017)
Ross MT, Flores D, Bertram R, Johnson F, Hyson RL. Neuronal Intrinsic Physiology Changes During Development of a Learned Behavior. eNeuro 2017 Oct 20; 4(5). pii: ENEURO.0297-17.2017. doi: 10.1523/ENEURO.0297-17.2017. PubMed PMID: 29062887 PubMed Link.
How is learning encoded by the neural network that controls birdsong? It has long been assumed that the intrinsic electrical properties of the component neurons are stable, with learning focused exclusively on synaptic plasticity within the network. Matthew Ross (Graduate Student, Hyson Lab) took the lead on this project to measure the intrinsic electrical properties of song network neurons as a function of the developmental stages of learning. A number of striking developmental changes in the intrinsic electrical properties of song network neurons were discovered, which were then converted to mathematical models by Diana Flores (Graduate Student, Bertram Lab) to deduce the specific ionic currents that correlate with song learning. The data show that the intrinsic electrical properties of song network neurons are not stable over the course of development and the observed plasticity that may have a role in encoding the learning of birdsong. The image below depicts the change in electrical excitability that occurs in one of the subpopulations of song network neurons as juvenile birds develop and learn their songs.
A Distributed Neural Network Model for the Distinct Roles of Medial and Lateral HVC in Zebra Finch Song Production (2017)
Galvis D, Wu W, Hyson RL, Johnson F, Bertram R. A Distributed Neural Network Model for the Distinct Roles of Medial and Lateral HVC in Zebra Finch Song Production. J Neurophysiol. 2017 Apr 5:jn.00917.2016. doi: 10.1152/jn.00917.2016. PubMed PMID: 28381490 PubMed Link.
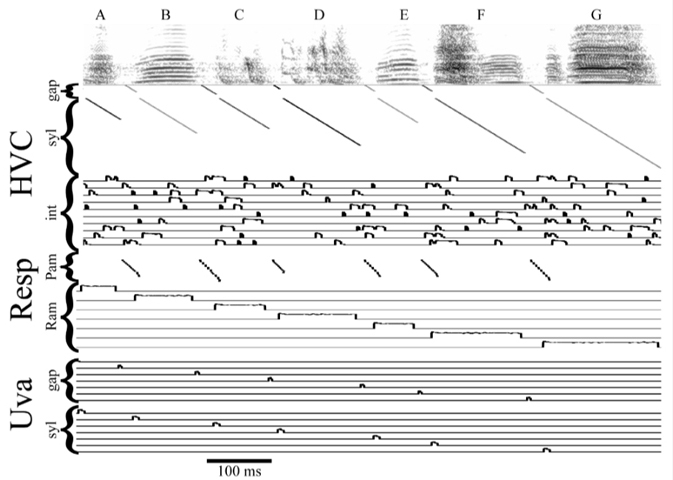
Danny Galvis (Graduate Student, Bertram Lab) took the lead on development of this biophysical model where individual syllables, the gaps of silence between syllables, and the syllable sequence are encoded by parallel streams of neural activity that traverse medial and lateral HVC. The structure of the model integrates neurobiological and behavioral data generated by Mark Basista and Kevin Elliott (Basista et al. 2014, Elliott et al. 2017). In the image below, we show a frequency spectrogram of the syllable sequence of a zebra finch song (top), the associated patterns of HVC and respiratory (Resp) activity that drive song, and the thalamic feedback to HVC (Uva) that signals the start of each syllable and gap of silence in the song.
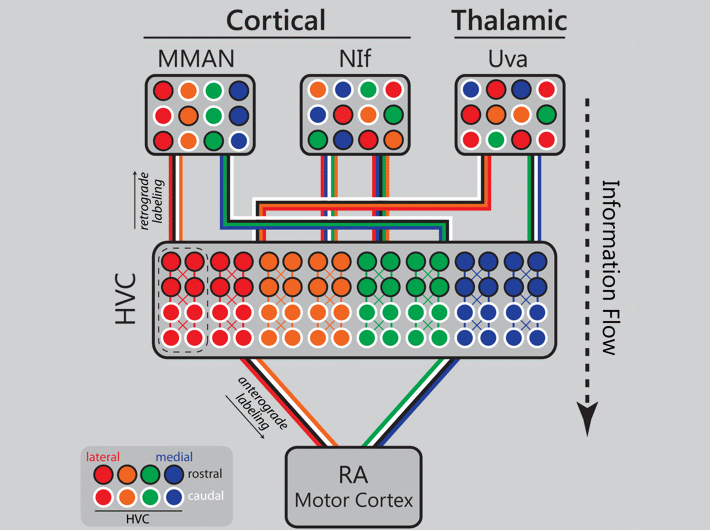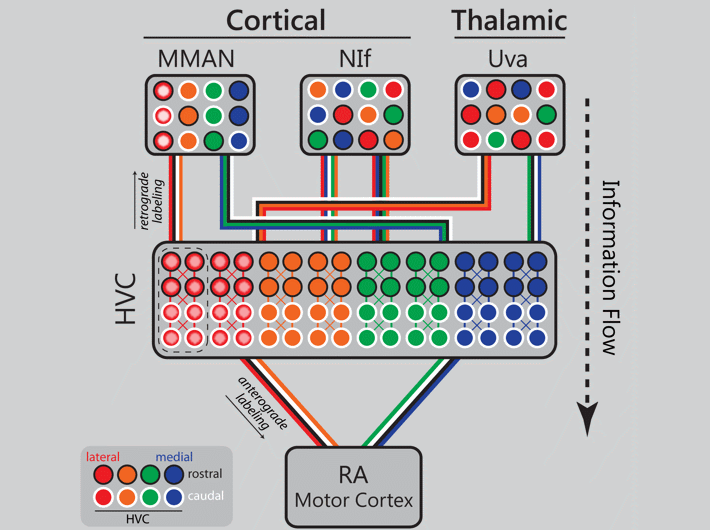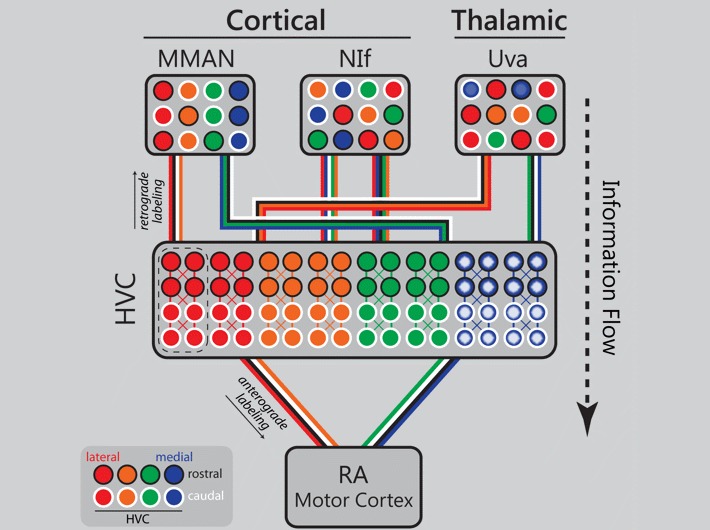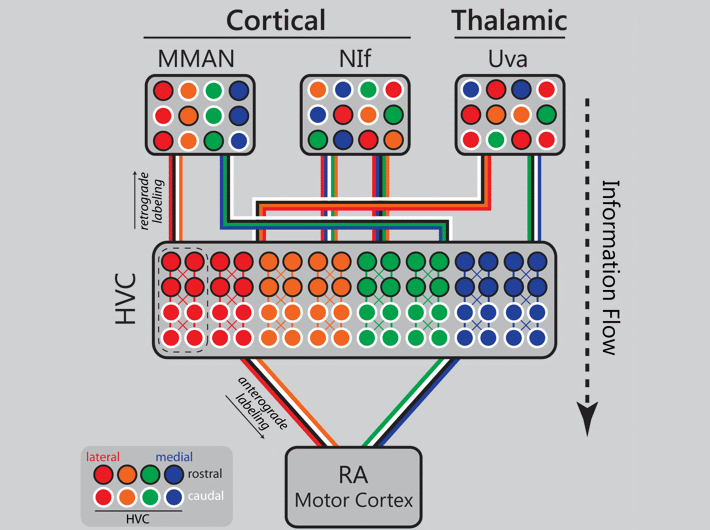
Orthogonal topography in the parallel input architecture of songbird HVC (2017)
Elliott KC, Wu W, Bertram R, Hyson RL, Johnson F. Orthogonal topography in the parallel input architecture of songbird HVC. J Comp Neurol. 2017 Jun 15;525(9):2133-2151. doi: 10.1002/cne.24189. Epub 2017 Mar 30. PubMed PMID: 28188629. PubMed Link.
Kevin Elliott (2016 Ph.D. graduate) took the lead on this project which demonstrates that the input architecture of HVC is massively parallel, and in some cases topographic. Danny Galvis used these experimental data to inform his network model of song encoding (Galvis et al. 2017). In the image below an animated gif highlights the distinct patterns of cortical and thalamic input received by HVC. Now imagine all three inputs active together on a millisecond time scale and you begin to get an idea of the intricate choreography of neural activity in a functioning brain.
Independent premotor encoding of the sequence and structure of birdsong in avian cortex (2014)
Basista MJ, Elliott KC, Wu W, Hyson RL, Bertram R, Johnson F. Independent premotor encoding of the sequence and structure of birdsong in avian cortex. J Neurosci. 2014 Dec 10;34(50):16821-34. doi: 10.1523/JNEUROSCI.1940-14.2014. PubMed PMID: 25505334. PubMed Link.
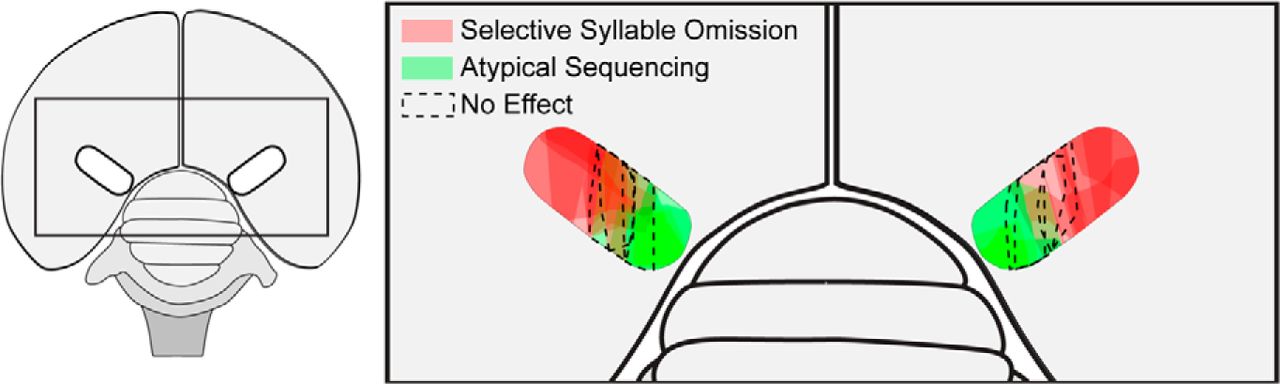
Mark Basista and Kevin Elliott (2016 Ph.D. graduates) took the lead on this project which demonstrated a functional topography across the lateral-medial axis of HVC. Mark's ablation data, complemented by Kevin's tract-tracing data, showed that lateral and medial HVC play independent roles in the encoding of song syllables and their sequence, respectively. Danny Galvis' used these experimental data to inform his network model of song encoding (Galvis et al. 2017). The image below shows an overhead view of the zebra finch brain and maps the behavioral effects of Mark's bilateral ablations of HVC. Ablations within the lateral portion of HVC induced the omission of song syllables (red shading) while ablations within the medial portion of HVC induced the atypical sequencing of song syllables (green shading). Interestingly, no effect was observed (dashed lines) when ablations targeted the border region between the two functional subdivisions of HVC.
Two neural streams, one voice: pathways for theme and variation in the songbird brain (2014)
Bertram R, Daou A, Hyson RL, Johnson F, Wu W. Two neural streams, one voice: pathways for theme and variation in the songbird brain. Neuroscience. 2014 Sep 26;277:806-17. doi: 10.1016/j.neuroscience.2014.07.061. Epub 2014 Aug 8. Review. PubMed PMID: 25106128. PubMed Link.
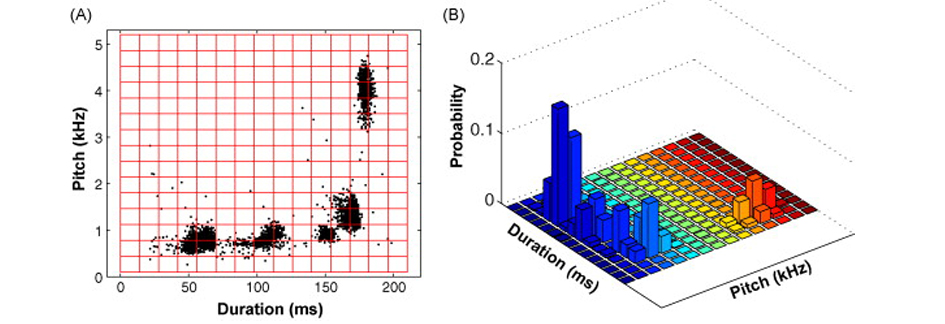
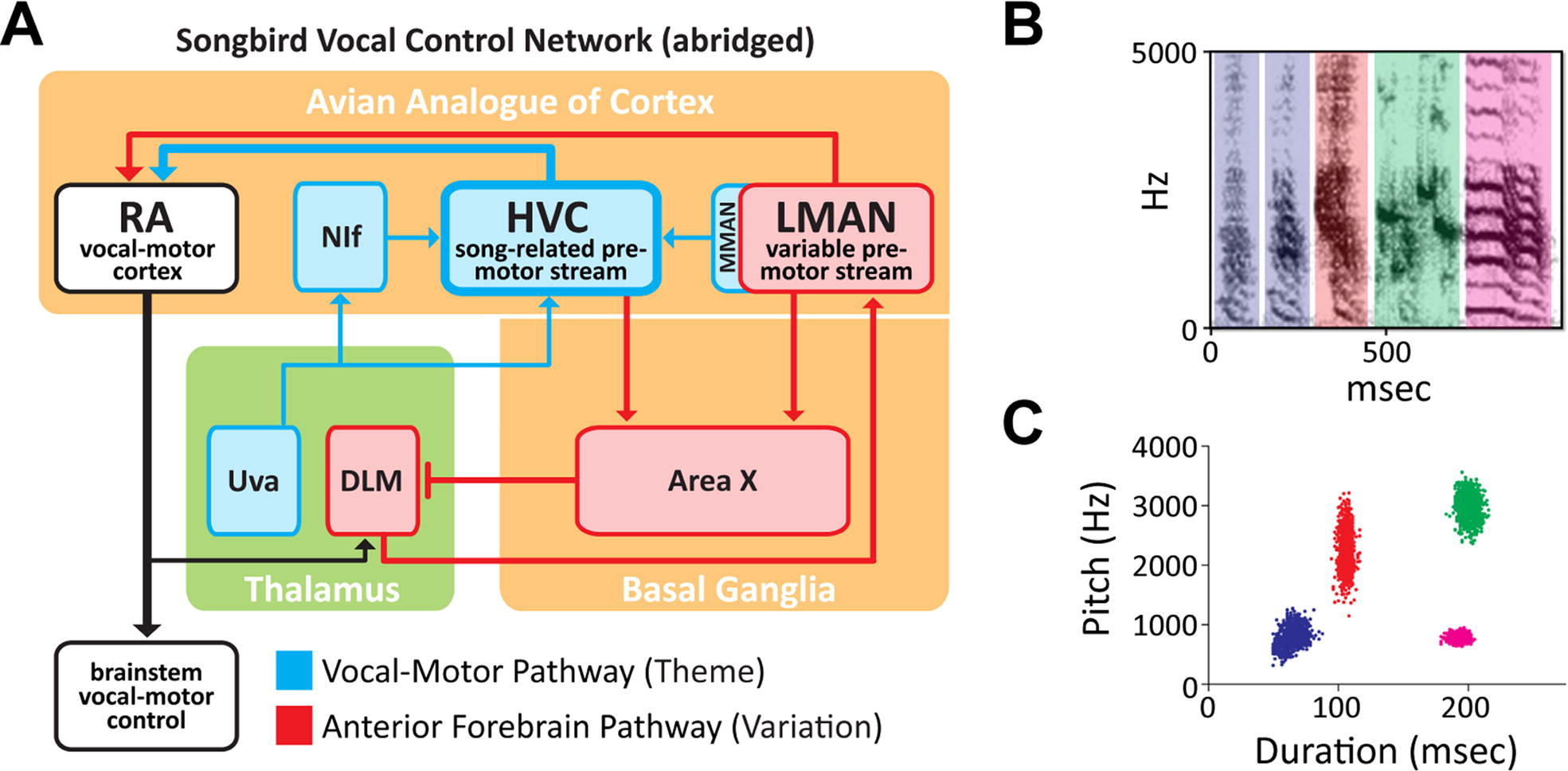
This review paper, authored by the faculty members of the Group and Arij Daou (2012 Ph.D. graduate), highlights the distinctive "dual premotor architecture" of the brain network that controls song. In the image below we show a sagittal schematic of the songbird vocal control network (A) where one pathway encodes the learned song (blue, "theme") while a second pathway encodes acoustic variation (red, "variation"). Both pathways converge at vocal-motor cortex (RA), where signals from the two pathways are integrated to drive song. As a result, the acoustic structure of the syllables that make up a male zebra finch song (see frequency spectrogram in B) varies over the course of a day of singing. The scatter plot (C) shows the duration and pitch of all syllables produced during a day of singing; each data point is one instance of a syllable and color coding of syllable clusters matches the spectrogram in (B). The clustering demonstrates the consistent “theme” of repeated syllables and the spread of each syllable cluster represents the “variation” that is apparent between repetitions of each syllable across the day of singing.
Disconnection of a basal ganglia circuit in juvenile songbirds attenuates the spectral differentiation of song syllables (2014)
Elliott KC, Wu W, Bertram R, Johnson F. Disconnection of a basal ganglia circuit in juvenile songbirds attenuates the spectral differentiation of song syllables. Dev Neurobiol. 2014 Jun;74(6):574-90. doi: 10.1002/dneu.22151. Epub 2013 Dec 14. Erratum in: Dev Neurobiol. 2014 Dec;74(12):1277. PubMed PMID: 24218118; PubMed Central PMCID: PMC4120835. PubMed Link.
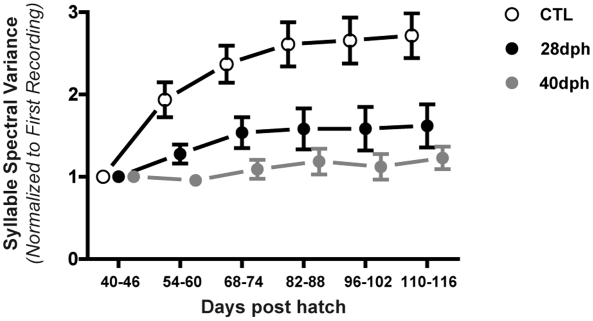
Kevin Elliott (2016 Ph.D. graduate) led this project to test the role of "babbling" in juvenile song learning. Like human infants, juvenile zebra finches begin the process of vocal learning with a period of babbling that is driven by a dedicated basal ganglia circuit. The adult song is subsequently learned and encoded by a separate cortical circuit. The image below shows the normal learning-related increase in syllable complexity (CTL) as juvenile birds transition from basal ganglia-driven babbling to cortical-driven adult song. However, the learning curve flattens in juvenile birds that were unable to babble due to surgical disconnection of the basal ganglia circuit (at 28 or 40 days post-hatch, dph). Kevin's findings show that the activity of the basal ganglia circuit - and the unstructured babbling that it generates - is essential for the cortical circuit to learn the complex and highly-structured songs of adults.
Electrophysiological characterization and computational models of HVC neurons in the zebra finch (2013)
Daou A, Ross MT, Johnson F, Hyson RL, Bertram R. Electrophysiological characterization and computational models of HVC neurons in the zebra finch. J Neurophysiol. 2013 Sep;110(5):1227-45. doi: 10.1152/jn.00162.2013. Epub 2013 May 29. PubMed PMID: 23719205. PubMed Link.
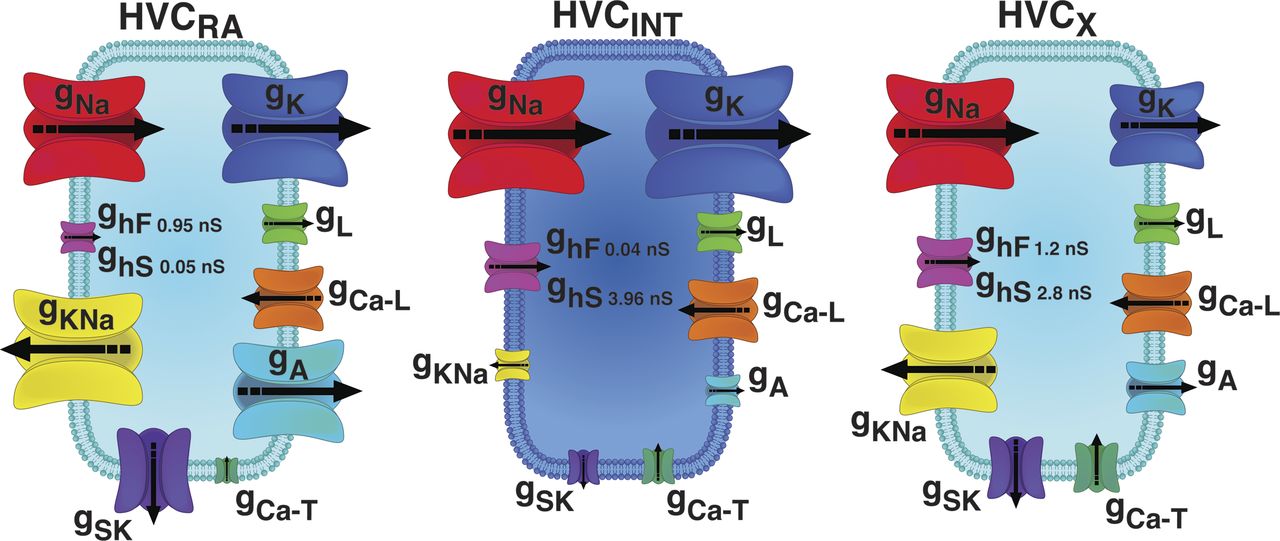
HVC is home to the neurons that encode the learned song of adult zebra finches. Arij Daou (2012 Ph.D. graduate) and Matthew Ross (Graduate Student, Hyson Lab) led this project to characterize the three major classes of HVC neurons - those that project to vocal-motor cortex (HVCRA), HVC interneurons (HVCINT) and HVC neurons that project to the basal ganglia (HVCX). Arij and Matthew measured the intrinsic physiology of each class of HVC neurons and then Arij developed biophysical mathematical models for each class. Model predictions about the role of specific ion currents in shaping the unique physiology of each class were then validated with pharmacological manipulation of different ion channels. These data were the first step towards developing a biophysically-valid computational model of song encoding within HVC. The image below shows a graphical depiction of the intrinsic physiology of each class of HVC neuron, where the size of ion channels has been log-scaled to reflect the direction and magnitude of electrical current carried by each type of ion channel - beautiful and informative. Software for mathematical models of each class of HVC neuron is freely available - get code.
A computational tool for automated large-scale analysis and measurement of bird-song syntax (2012)
Daou A, Johnson F, Wu W, Bertram R. A computational tool for automated large-scale analysis and measurement of bird-song syntax. J Neurosci Methods. 2012 Sep 30;210(2):147-60. doi: 10.1016/j.jneumeth.2012.07.020. Epub 2012 Aug 6. PubMed PMID: 22890237. PubMed Link.
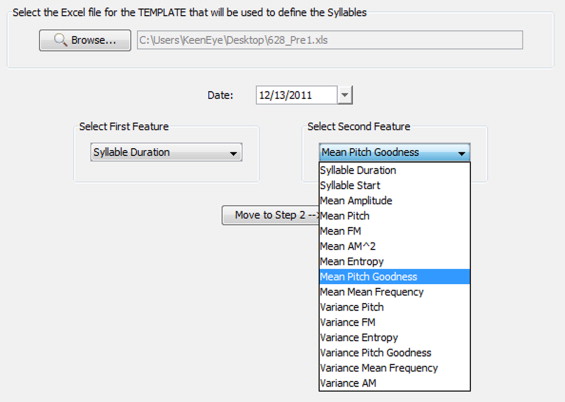
Arij Daou (2012 Ph.D. graduate) led this project to develop a Java-based software program to automate the identification of specific syllables and the measurement of syllable sequence (syntax). The software builds on and extends the capability of Sound Analysis Pro, the leading birdsong analysis tool. The image below shows one the dialog boxes that a user encounters, where specific acoustic features are used to identify particular song syllables. This software program is freely available - get code.
Axial organization of a brain region that sequences a learned pattern of behavior (2012)
Stauffer TR, Elliott KC, Ross MT, Basista MJ, Hyson RL, Johnson F. Axial organization of a brain region that sequences a learned pattern of behavior. J Neurosci. 2012 Jul 4;32(27):9312-22. doi: 10.1523/JNEUROSCI.0978-12.2012. PubMed PMID: 22764238; PubMed Central PMCID: PMC3400431. PubMed Link.
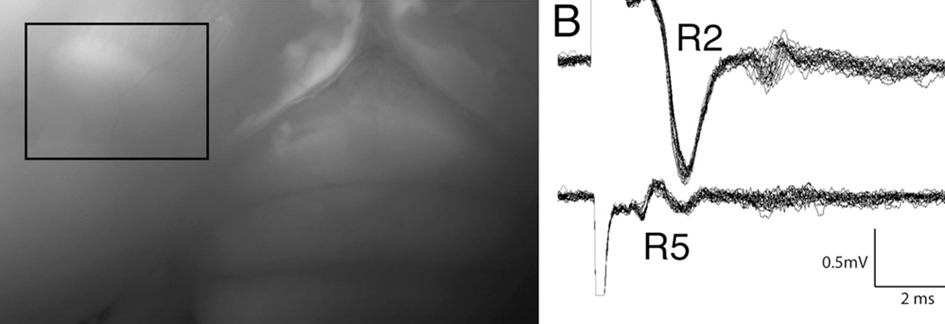
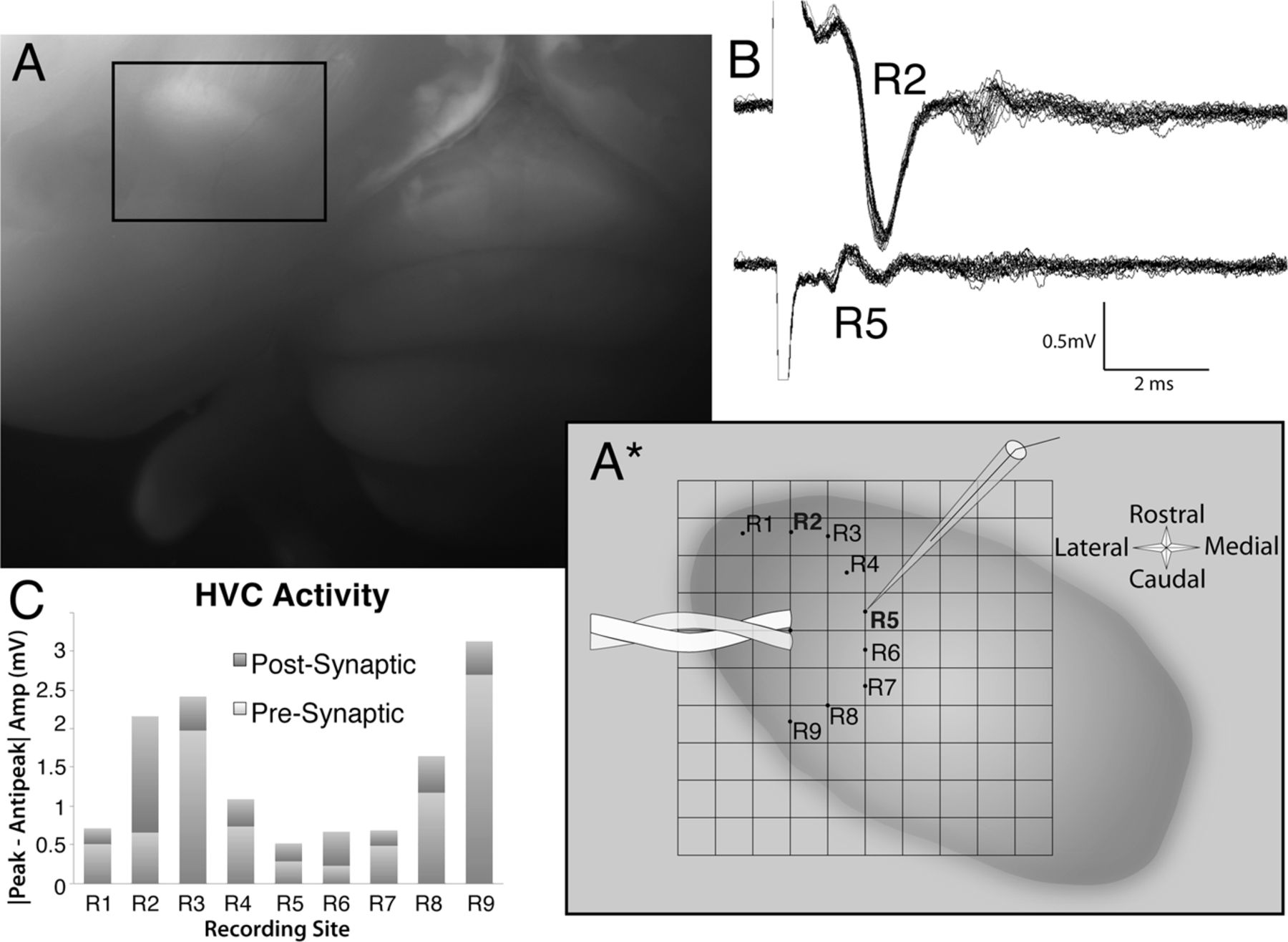
This paper marks the beginning of a general theme in the Group's work - HVC is far more organized in connectivity and function than previously believed. Tiffanie Stauffer (2010 Ph.D. graduate) led this project, where she used an activity marker to demonstrate that HVC neurons are organized into multiple parallel "swaths" that are connected to one another primarily within the rostral-caudal axis. This remarkable result was supported by tract-tracing (Kevin Elliott, 2016 Ph.D. graduate) and electrophysiological data (Matthew Ross, Graduate Student, Hyson Lab, and Mark Basista, 2016 Ph.D. graduate). The image below shows some of the stimulation and recording results from Matthew and Mark's work, where stimulation within the central portion of HVC elicited robust, synchronous responses when recording electrodes were placed within the rostral-caudal axis (R2, R3, R8, R9), but not when they were placed within the medial-lateral axis (R4, R5, R6, R7).
Dual premotor contribution to songbird syllable variation (2011)
Thompson JA, Basista MJ, Wu W, Bertram R, Johnson F. Dual pre-motor contribution to songbird syllable variation. J Neurosci. 2011 Jan 5;31(1):322-30. doi: 10.1523/JNEUROSCI.5967-09.2011. PubMed PMID: 21209218; PubMed Central PMCID: PMC3080041. PubMed Link.
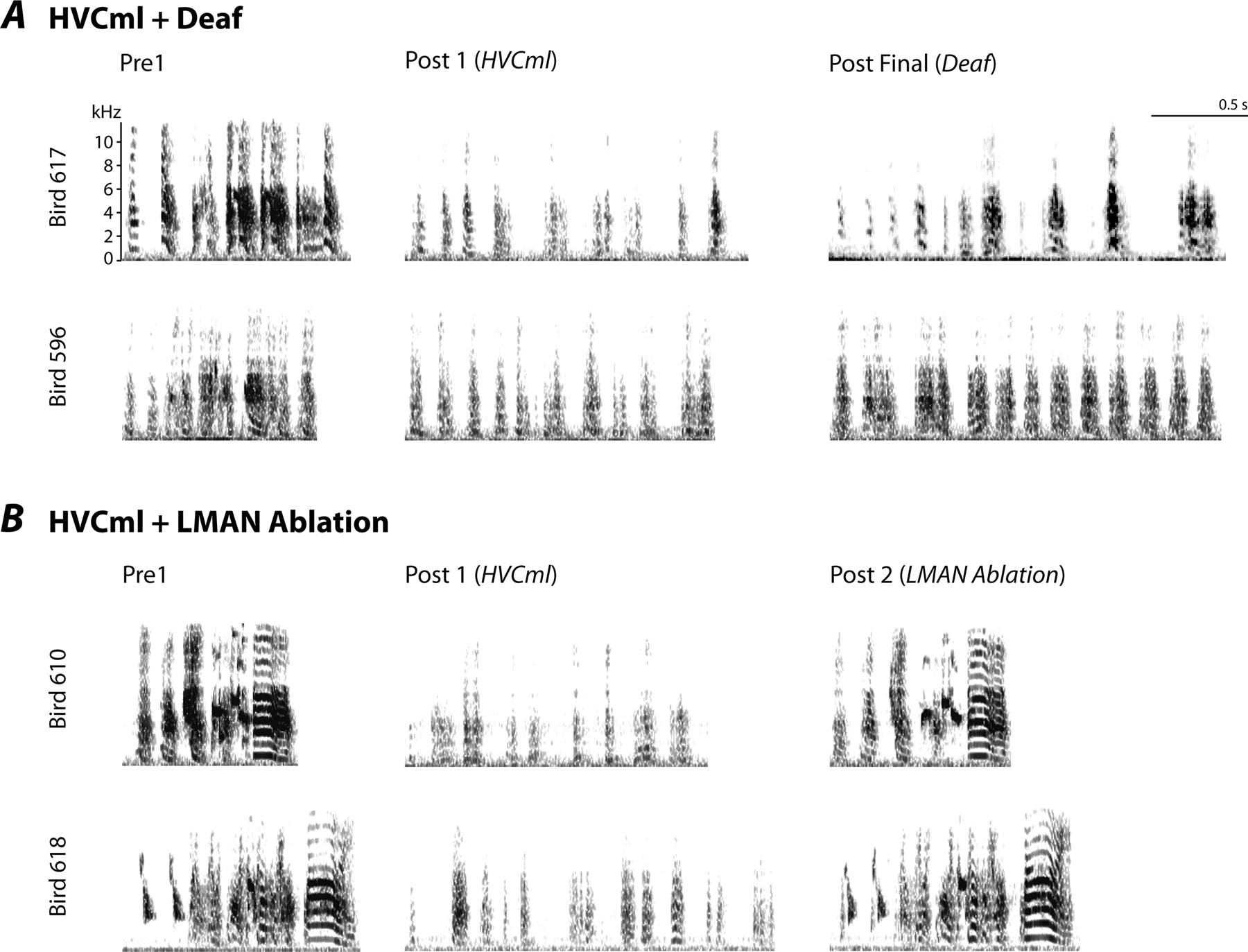
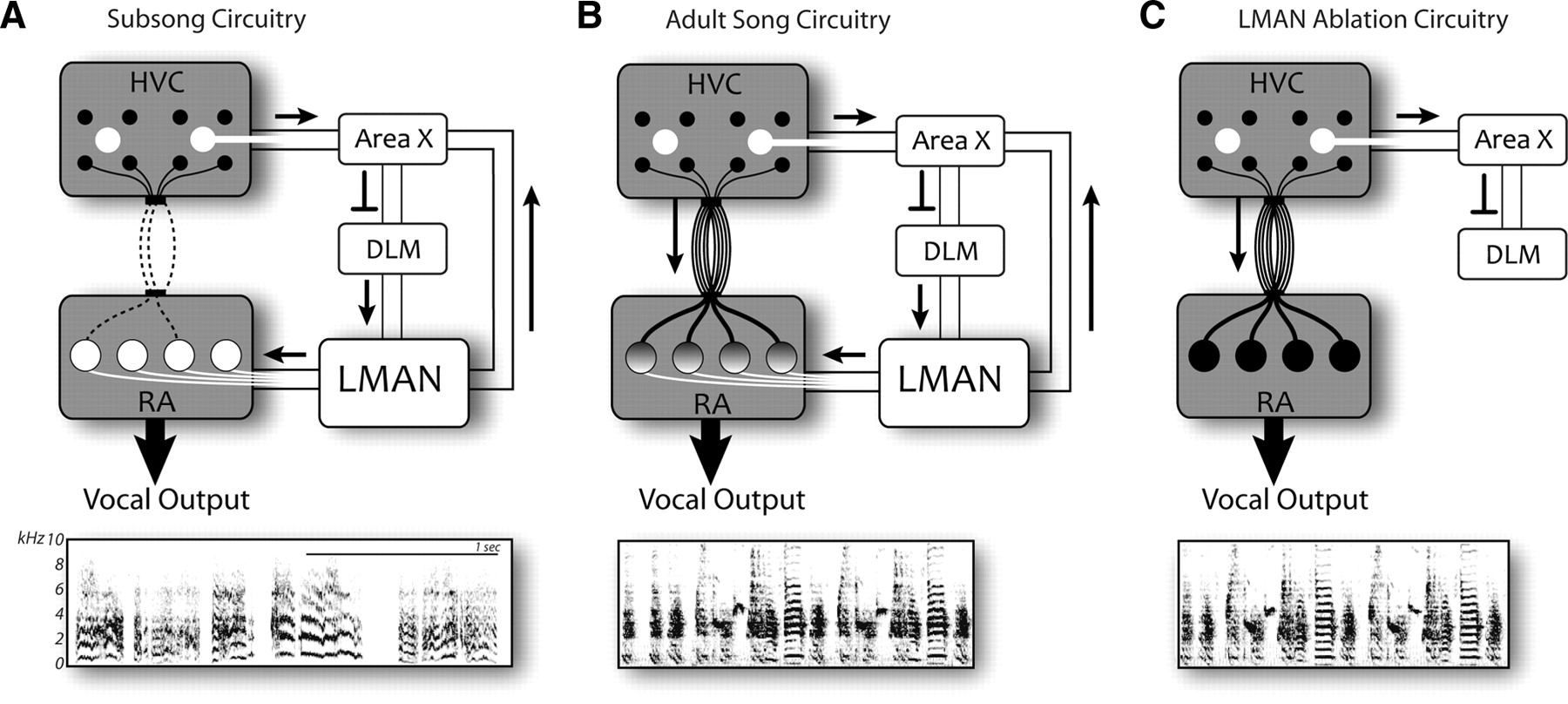
John Thompson (2008 Ph.D. graduate) and Mark Basista (2016 Ph.D. graduate) led this project to identify premotor sources of spectral variation in the production of song syllables. The image below shows schematics of the connectivity of cortical (grey) and basal ganglia (white) circuits in juveniles just beginning the process of song learning (A), adult birds producing learned song (B), and adult birds following disconnection of the basal ganglia circuit (LMAN ablation, C). Below each schematic is an example of the vocalizations produced in each circuit configuration. During juvenile subsong (A), the basal ganglia circuit dominates vocal output, driving the vocal organ to produce highly unstructured variation in the acoustic properties of song syllables. In adult birds (B), the cortical circuit dominates vocal output, driving the vocal organ to produce the highly structured acoustic variation of the learned vocal pattern. However, the basal ganglia circuit still contributes modest spectral and temporal jitter to the ongoing production of adult song. Disconnection of the basal ganglia circuit in adult birds (C) results in a more acoustically pure version of the learned song - the reduction in noise and the increase in tonal quality is apparent by comparing the frequency spectrograms in B and C.
A statistical method for quantifying songbird phonology and syntax (2008)
Wu W, Thompson JA, Bertram R, Johnson F. A statistical method for quantifying songbird phonology and syntax. J Neurosci Methods. 2008 Sep 15;174(1):147-54. doi: 10.1016/j.jneumeth.2008.06.033. Epub 2008 Jul 11. PubMed PMID: 18674560; PubMed Central PMCID: PMC2569874. PubMed Link.
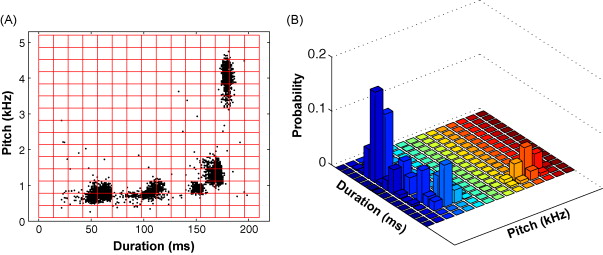
Wei Wu and John Thompson (2008 Ph.D. graduate) led this project to develop a new statistical method to detect significant shifts in the acoustic structure of song. The method, inspired by Shannon's Information Theory, enables statistical comparison across entire days of singing, effectively removing sampling bias and the effect of outliers on estimates of central tendency and statistical dispersion. The image below shows a duration x pitch scatterplot of a day of singing from an example bird (A) where each data point is one instance of a syllable. Clusters of data points reflect repeated instances of a particular syllable. To detect change in the acoustic structure of song by Kullback-Leibler distance (an information-theoretic quantity with units in bits) each day of singing is converted to a probability density distribution (B). The software for this analysis is freely available - get code.
Auditory-dependent vocal recovery in adult male zebra finches is facilitated by lesion of a forebrain pathway that includes the basal ganglia (2007)
Thompson JA, Wu W, Bertram R, Johnson F. Auditory-dependent vocal recovery in adult male zebra finches is facilitated by lesion of a forebrain pathway that includes the basal ganglia. J Neurosci. 2007 Nov 7;27(45):12308-20. PubMed PMID: 17989295. PubMed Link.
John Thompson (2008 Ph.D. graduate) led this project to understand why partial ablation (microlesion) of HVC caused an immediate dissolution of adult song, followed within 1-2 weeks by complete recovery. In a first experiment, John demonstrated that deafened birds could not recover their songs following HVC microlesions, indicating that recovery of song depends on auditory feedback (A in the image below). The auditory-dependence of the gradual recovery from HVC microlesions suggests that auditory feedback may restore song by rebalancing the strength of the dual premotor streams to favor HVC. If so, John reasoned that he should be able to induce a sudden recovery of song if he followed HVC microlesions with ablation of the output of a basal ganglia circuit (LMAN), which is what happened (B in the image below). This surprising result - that the behavioral effects of injury to one part of the brain (HVC) could be immediately resolved by removal of another part of the brain (LMAN) - was a key data point in elucidating the dual premotor control of song in zebra finches. It is now understood that HVC microlesions merely weaken HVC signaling, indirectly amplifying the variable premotor activity of the basal ganglia circuit, which leads to unstructured singing. Disconnection of the basal ganglia circuit restores the primacy of HVC signaling in controlling vocal output, and the structure of the learned adult song.